Dimorphism and mating in Ascomycetes,
Zygomycetes and Basidiomycetes
Zygomycetes and Basidiomycetes
Purpose
The objectives of this laboratory exercise on September 26, 2012, were to 1) microscopically observe the three spore types (ascospores, macroconidia, microconidia) of Neurospora crassa 2) cross strains of N. crassa with different mating type alleles in vitro, and confirm or determine mating type alleles using a GFP-labeled strain 3) observe dimorphism (hyphae and yeast) in Mucor rouxii 4) learn about the life cycle of Ustilago maydis that causes common corn smut and observe characteristic symptom development in two inoculated commercial corn cultivars.
Materials and Methods
Prepared cultures of N. crassa and M. rouxii
Prepared spore suspension of U. maydis haploid strain Sg200
Inoculated (Silver Queen and OPR2-1) and non-inoculated (OPR2-1) corn seedlings
Westergaard and Mitchell synthetic crossing (SC) solid medium
Microscope slides
Cover slips
Scalpel
Dissecting needle
Needle and syringe
Dropper bottle with water
Sterile deionized water
Spray bottle with 70% ethanol
Parafilm
Permanent marker
Paper towels
Metal striker
Bunsen burner
Canon PowerShot SD550 digital camera
Olympus CX31 compound microscope
Prepared cultures of N. crassa and M. rouxii
Prepared spore suspension of U. maydis haploid strain Sg200
Inoculated (Silver Queen and OPR2-1) and non-inoculated (OPR2-1) corn seedlings
Westergaard and Mitchell synthetic crossing (SC) solid medium
Microscope slides
Cover slips
Scalpel
Dissecting needle
Needle and syringe
Dropper bottle with water
Sterile deionized water
Spray bottle with 70% ethanol
Parafilm
Permanent marker
Paper towels
Metal striker
Bunsen burner
Canon PowerShot SD550 digital camera
Olympus CX31 compound microscope
A. Observation of N. crassa spore types
Using a sterile technique (described in the report from the laboratory on September 5, 2012, that is posted in my blog), a dissecting needle was used to remove mycelia containing conidiophores and spores growing on the inside wall of a disposable 50-ml centrifuge tube. The needle was touched to a drop of water on a microscope slide. A cover slip was applied, and then the sample was viewed with a compound microscope that had been set to Köhler (described in the report from the laboratory on September 5, 2012, that is posted in my blog). Again, using a sterile technique, ~1-2 mm plugs were cut, using a scalpel, from cultures on nutrient-poor and nutrient-rich media prepared prior to the laboratory exercise, and then the plugs were placed in drops of water on microscope slides. To avoid disrupting the configuration of fungal structures, cover slips were applied but were not pressed to squash the specimens prior to viewing with the same compound microscope.Photographs of fungal structures were taken through the eyepiece of the microscope using a Canon PowerShot SD550 digital camera.
Using a sterile technique (described in the report from the laboratory on September 5, 2012, that is posted in my blog), a dissecting needle was used to remove mycelia containing conidiophores and spores growing on the inside wall of a disposable 50-ml centrifuge tube. The needle was touched to a drop of water on a microscope slide. A cover slip was applied, and then the sample was viewed with a compound microscope that had been set to Köhler (described in the report from the laboratory on September 5, 2012, that is posted in my blog). Again, using a sterile technique, ~1-2 mm plugs were cut, using a scalpel, from cultures on nutrient-poor and nutrient-rich media prepared prior to the laboratory exercise, and then the plugs were placed in drops of water on microscope slides. To avoid disrupting the configuration of fungal structures, cover slips were applied but were not pressed to squash the specimens prior to viewing with the same compound microscope.Photographs of fungal structures were taken through the eyepiece of the microscope using a Canon PowerShot SD550 digital camera.
B. In vitro crossing of N. crassa strains with different mating types alleles
Using a permanent marker, four disposable Petri dishes containing nutrient-poor SC solid medium were divided in half on the underside, and each half was labeled with a strain of N. crassa in which the mating type allele is either known or not known. Using a sterile technique, a dissecting needle was used to remove mycelia containing conidiophores and spores of a strain of N. crassa growing on the inside wall of a micro-culture tube. The needle then was touched to the solid medium at the center outer edge of the half designated for the strain. Inoculated dishes were sealed with parafilm and incubated at 25 degrees Celsius. Paired strains were observed microscopically during the laboratory exercise on October 3 for the presence or absence of hyphal fusion, as evidenced by the formation of perithecia, and to confirm or determine mating type alleles. Photographs of pairings were taken using a Canon PowerShot SD550 digital camera.
Paired N. crassa strains are as follows:
SMRP10 (A type) x SMRP11 (a type)
Using a permanent marker, four disposable Petri dishes containing nutrient-poor SC solid medium were divided in half on the underside, and each half was labeled with a strain of N. crassa in which the mating type allele is either known or not known. Using a sterile technique, a dissecting needle was used to remove mycelia containing conidiophores and spores of a strain of N. crassa growing on the inside wall of a micro-culture tube. The needle then was touched to the solid medium at the center outer edge of the half designated for the strain. Inoculated dishes were sealed with parafilm and incubated at 25 degrees Celsius. Paired strains were observed microscopically during the laboratory exercise on October 3 for the presence or absence of hyphal fusion, as evidenced by the formation of perithecia, and to confirm or determine mating type alleles. Photographs of pairings were taken using a Canon PowerShot SD550 digital camera.
Paired N. crassa strains are as follows:
SMRP10 (A type) x SMRP11 (a type)
SMRP10 (A type) x NCAL (unknown type)
SMRP10 (A type) x CSP-1 GFP (unknown type)
SMRP11 (a type) x CSP-1 GFP (unknown type)
C. Observation of M. rouxii dimorphism
SMRP11 (a type) x CSP-1 GFP (unknown type)
C. Observation of M. rouxii dimorphism
 |
| Using a scalpel under sterile conditions, a slice was removed from an agar plug inoculated prior to the laboratory exercise with M. rouxii. |
 |
| The slice of agar plug inoculated with M. rouxii was mounted on a slide for microscopic viewing of hyphae and yeast cells produced by the dimorphic fungus. The top surface of the slice is facing toward the viewer. |
D. Inoculation of commercial corn cultivar seedlings with U. maydis strain Sg200 and observance of common corn smut symptom development
A sterile needle and syringe was used to inoculate one OPR2-1 commercial corn cultivar seedling with sterile deionized water (control plant) and another with a spore suspension of U. maydis haploid strain Sg200 prepared prior to the laboratory exercise. The control and inoculated plants were placed in a rack under fluorescent grow lights and watered regularly for the duration of the experiment. Both plants were macroscopically observed 7, 10 and 14 days post-inoculation for symptoms characteristic of a successful common corn smut infection.
Seedlings of commercial corn cultivars Silver Queen and OPR2-1 inoculated with the same strain of U. maydis 7 and 13 days, respectively, prior to the start of the laboratory exercise also were macroscopically observed for an additional two weeks, and any symptom development was compared with any symptom development on plants inoculated during the laboratory. Photographs of of inoculated plants were taken using a Canon PowerShot SD550 digital camera.
A sterile needle and syringe was used to inoculate one OPR2-1 commercial corn cultivar seedling with sterile deionized water (control plant) and another with a spore suspension of U. maydis haploid strain Sg200 prepared prior to the laboratory exercise. The control and inoculated plants were placed in a rack under fluorescent grow lights and watered regularly for the duration of the experiment. Both plants were macroscopically observed 7, 10 and 14 days post-inoculation for symptoms characteristic of a successful common corn smut infection.
Seedlings of commercial corn cultivars Silver Queen and OPR2-1 inoculated with the same strain of U. maydis 7 and 13 days, respectively, prior to the start of the laboratory exercise also were macroscopically observed for an additional two weeks, and any symptom development was compared with any symptom development on plants inoculated during the laboratory. Photographs of of inoculated plants were taken using a Canon PowerShot SD550 digital camera.
Results
A. Observation of N. crassa spore types
 |
| Ascospore from a culture prepared in a disposable 50-ml centrifuge tube (40X). Specimen removed from culture using a dissecting needle. Photograph cropped and enlarged to show detail. |
 |
| Chains of macroconidia and conidiophores from a culture prepared on nutrient-rich medium (40X). Specimen removed from culture using a dissecting needle. Photograph cropped and enlarged to show detail. |
 |
| Developing chains of macroconidia from a culture prepared on nutrient-rich medium (40X). Specimen removed from culture using a scalpel. Photograph cropped and enlarged to show detail. |
 |
| Developing chains of macroconidia from a culture prepared on nutrient-rich medium (40X). Specimen removed from culture using a scalpel. Photograph cropped and enlarged to show detail. |
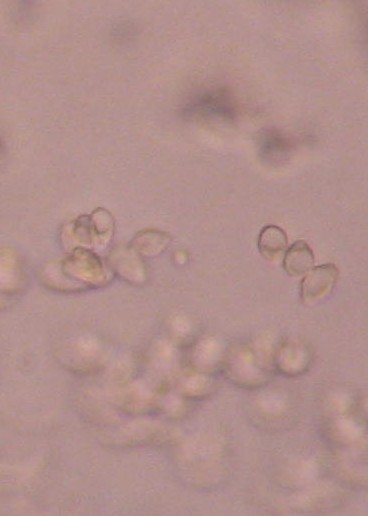 |
| Microconidia (smaller, round object in center of photograph) from a culture prepared on nutrient-poor SC medium (40X). Specimen removed from culture using a scalpel. Photograph cropped and enlarged to show detail. |
B. In vitro crossing of N. crassa strains with different mating type alleles
SMRP10 x SMRP11
SMRP10 x NCAL
Pairing is successful, as evidenced by the presence of perithecia on both sides of the plate.
 |
| Top of plate |
 |
| Bottom of plate |
My plate is incognito, so I do not have results to post from this pairing.
SMRP10 x CSP-1 GFP
SMRP11 x CSP-1 GFP
Pairing is successful, as evidenced by a line of perithecia between the two strains. CSP-1 GFP appears to be mating type "a" as SMRP10 is mating type "A."
 |
| Top of plate. |
 |
| Bottom of plate |
 |
| Perithecia |
Pairing is not successful, as evidenced by a lack of perithecia. Results would be expected, as SMRP11 and CSP-1 GFP are both mating type "a."
 |
| Top of plate |
{kind=link}
 |
| Bottom of plate |
C. Observation of M. rouxii dimorphism
 | |||
| Yeast cells observed ~8mm below the top surface of a vertical slice from an inoculated agar plug (40X). Photograph cropped and enlarged to show detail. |
D. Common corn smut symptom development in Silver Queen and OPR2-1 commercial corn cultivar seedlings inoculated with U. maydis strain Sg200
I like the following figures detailing the life cycle of U. maydis, so I included both in this blog.

I like the following figures detailing the life cycle of U. maydis, so I included both in this blog.
http://www.metapathogen.com/IMG/ustilago-life-cycle.png
 | |||||
| Image from Steinberg and
Perez-Martin. 2008. Trends Cell Biol. 18:61. http://www.gerosteinberg.com/images/modelfig1.gif The following reference also discusses the morphogenesis of U. maydis in corn during teliospore formation. https://docs.google.com/file/d/0BzR22NlmXyPVeS1CS0JKTTJVSUk/edit?pli=1 |
 |
OPR2-1 commercial corn cultivar 0 days post inoculation with U. maydis haploid strain Sg200 that causes common corn smut (top photo). The control plant (top, right) was inoculated with sterile water. The plant inoculated with U. maydis showed symptoms of infection after 7 days (bottom photo). Notice the appearance of small yellow tumors along the midrib of the leaf blade and a bluish discoloration of leaf tissue indicative of induction of anthocyanin. Anthocyanins are water-soluble vacuolar pigments that also may appear red or purple, depending on pH. They belong to a parent class of molecules called flavonoids that are synthesized via the phenylpropanoid pathway and occur in all tissues of higher plants..jpg) |
.jpg) |
| Common corn smut symptoms on a leaf of commercial corn cultivar OPR2-1 inoculated with U. maydis haploid strain Sg200. This seedling was inoculated 13 days prior to the start of the current laboratory exercise. The photograph was taken during the current laboratory exercise. Note the reddish anthocyanin streaking. Tumors were not observed on this plant. Symptoms of a possible infection did not progress during the next 7 days of observation. |
+%232.jpg) |
| Common smut symptoms on a leaf of commercial corn cultivar Silver Queen inoculated with U. maydis haploid strain Sg200. This seedling was inoculated 7 days prior to the start of the current laboratory exercise. The photograph was taken during the current laboratory exercise. Note the reddish anthocyanin streaking and formation of yellow tumors indicative of a successful infection, which appears to have occurred sooner and possibly more severely on this cultivar than on the OPR2-1 cultivar. However, the disease failed to progress during the next 7 days of observation. |
Discussion
Macroconidia of N. crassa were more abundant and thus more readily observed microscopically. Only one ascospore of the Ascomycetous fungus was observed microscopically. Using a scalpel to remove a small plug from a plated culture works better than a dissecting needle. The scalpel makes sharper cuts and can be used to get under and lift the plug. The plug is essentially destroyed along with the fungal structures when using a dissecting needle. However, the dissecting needle is perfect for removing mycelia containing conidiophores and spores from culture tubes.
Yeast cells but no hyphae were observed in agar plugs inoculated prior to the laboratory exercise with M. rouxii. Thus, the fungus was given more time to develop so that dimorphism in the Zygomycete may be observed. Plugs were scheduled to be reexamined during the laboratory exercise on October 3, 2012. However, they had become contaminated with N. crassa during the week between the two labs. This was a prime example of the need to be meticulous in terms of sterility in the lab.
Preliminary observations of common corn smut symptoms on commercial corn cultivars inoculated prior to the start of the current laboratory exercise seemed to indicate that Silver Queen is more susceptible to U. maydis strain Sg200 than OPR2-1. Symptom development in the inoculated seedlings (yellow tumor formation) was indicative that a "dimorphic switch" occurred from the production of non-pathogenic, saprotrophic yeast-like sporidia to the production of conjugative, dikaryotic hyphae, which is necessary for successful infection by the Basidiomycete.
Disease development in my inoculated plant and in the plants inoculated prior to the start of this laboratory exercise was poor during the week following this lab. Thus, I waited another six days (until Oct. 9) before starting the process of removing the chloryphyll from the diseased leaf blade on my inoculated plant (seen in the above photo) with a 2:1 ethanol:acetic acid solution and then staining the leaf with 0.1% trypan blue plus lactophenol in order to microscopically observe any fungal structures within the leaf tissue. So I will not have pictures from that process until after the midterm grade. It is not known why infection failed to progress or not occur in some of the plants. It could have been due to the inoculum or the means of delivery to the plant. Plants were inoculated by injecting a spore suspension into the main stems using a needle and syringe. As an alternative, the spore suspension could have been sprayed onto plant leaves. A small agar plug from a pure culture of U. maydis also may have been placed on leaves or in the whorl of the main stem. That would make sense, as the fungus invades host epidermal and intracellular cells via hyphae, which then fragment to form tumors that in turn form teliospores.
Macroconidia of N. crassa were more abundant and thus more readily observed microscopically. Only one ascospore of the Ascomycetous fungus was observed microscopically. Using a scalpel to remove a small plug from a plated culture works better than a dissecting needle. The scalpel makes sharper cuts and can be used to get under and lift the plug. The plug is essentially destroyed along with the fungal structures when using a dissecting needle. However, the dissecting needle is perfect for removing mycelia containing conidiophores and spores from culture tubes.
Yeast cells but no hyphae were observed in agar plugs inoculated prior to the laboratory exercise with M. rouxii. Thus, the fungus was given more time to develop so that dimorphism in the Zygomycete may be observed. Plugs were scheduled to be reexamined during the laboratory exercise on October 3, 2012. However, they had become contaminated with N. crassa during the week between the two labs. This was a prime example of the need to be meticulous in terms of sterility in the lab.
Preliminary observations of common corn smut symptoms on commercial corn cultivars inoculated prior to the start of the current laboratory exercise seemed to indicate that Silver Queen is more susceptible to U. maydis strain Sg200 than OPR2-1. Symptom development in the inoculated seedlings (yellow tumor formation) was indicative that a "dimorphic switch" occurred from the production of non-pathogenic, saprotrophic yeast-like sporidia to the production of conjugative, dikaryotic hyphae, which is necessary for successful infection by the Basidiomycete.
Disease development in my inoculated plant and in the plants inoculated prior to the start of this laboratory exercise was poor during the week following this lab. Thus, I waited another six days (until Oct. 9) before starting the process of removing the chloryphyll from the diseased leaf blade on my inoculated plant (seen in the above photo) with a 2:1 ethanol:acetic acid solution and then staining the leaf with 0.1% trypan blue plus lactophenol in order to microscopically observe any fungal structures within the leaf tissue. So I will not have pictures from that process until after the midterm grade. It is not known why infection failed to progress or not occur in some of the plants. It could have been due to the inoculum or the means of delivery to the plant. Plants were inoculated by injecting a spore suspension into the main stems using a needle and syringe. As an alternative, the spore suspension could have been sprayed onto plant leaves. A small agar plug from a pure culture of U. maydis also may have been placed on leaves or in the whorl of the main stem. That would make sense, as the fungus invades host epidermal and intracellular cells via hyphae, which then fragment to form tumors that in turn form teliospores.
great job. Really informative. Thanks.
ReplyDelete