Fluorescence microscopy and protein localization; continuation of Neurospora genetics (screening progeny)
Purpose
The purpose of this laboratory exercise on October 24, 2012, was to 1) screen progeny from Neurospora crassa crosses initiated during Lab 5 on September 26, 2012, by picking individual mature ascospores and inoculating slants containing Vogel's minimal medium, 2) observe localization of green fluorescent protein (GFP) fusions with nuclei, actin, tubulin and Mak-2 in growing hyphae of N. crassa strains using fluorescence microscopy, 3) learn about a Fungal Genetics Stock Center website, and 4) observe basidia, basidiospores, clamp connections and Buller's drop in a field-collected mushroom (Chlorophyllum sp.).
Materials and Methods
A scalpel was used to cut a block from prepared 4% water agar (WA) medium (Figure 1 below). The block was placed on a microscope slide. A flame-sterilized metal inoculation loop was dipped in sterile water and then used to transfer ascospores from the culture dish lid of the N. crassa SMRP10 x CSP-1-GFP cross (refer to Labs 5 and 8 in my blog) to the WA block. The cross had been incubated for a week under continuous fluorescent lighting to facilitate maturation of perithecia and release of ascospores. A flame-sterilized platinum-iridium spore picker was then used (while viewing with a dissecting microscope) to cut an ~ 0.2 mm square containing an individual ascospore from the WA block and transfer it to a culture tube containing Vogel's minimal medium (Figure 2 below). A total of four ascospores were transferred to slants. The inoculated slants were placed overnight in a 4 degrees Celsius incubator to allow the spores to hydrate. The slants were then placed in a 65 degrees Celsius water bath for 45 minutes (heat shock) to kill any vegetative tissue and permit germination of the spores.
Results
B. Fluorescence microscopy of GFP localizations
D. Observation of fungal structures of Chlorophyllum sp.
N. crassa SMRP10 x CSP-1-GFP
Chlorophyllum sp. mushroom
Prepared slants of Vogel's minimal medium
Prepared 4% water agar medium
Platinum-iridium spore picker
Scalpel
Metal inoculation loop
Microscope slides
Microscope slides
Cover slips
Sterile water
Dropper bottle with water
Dropper bottle with water
Bunsen burner
Metal striker
4 degrees Celsius incubator
65 degrees Celsius water bath
Metal striker
4 degrees Celsius incubator
65 degrees Celsius water bath
Canon PowerShot SD550 digital camera
Olympus SZ30 zoom stereo microscope
Olympus CX31 compound microscope
Olympus SZ30 zoom stereo microscope
Olympus CX31 compound microscope
Fluorescence microscope
A. Screening of Neurospora progeny
A scalpel was used to cut a block from prepared 4% water agar (WA) medium (Figure 1 below). The block was placed on a microscope slide. A flame-sterilized metal inoculation loop was dipped in sterile water and then used to transfer ascospores from the culture dish lid of the N. crassa SMRP10 x CSP-1-GFP cross (refer to Labs 5 and 8 in my blog) to the WA block. The cross had been incubated for a week under continuous fluorescent lighting to facilitate maturation of perithecia and release of ascospores. A flame-sterilized platinum-iridium spore picker was then used (while viewing with a dissecting microscope) to cut an ~ 0.2 mm square containing an individual ascospore from the WA block and transfer it to a culture tube containing Vogel's minimal medium (Figure 2 below). A total of four ascospores were transferred to slants. The inoculated slants were placed overnight in a 4 degrees Celsius incubator to allow the spores to hydrate. The slants were then placed in a 65 degrees Celsius water bath for 45 minutes (heat shock) to kill any vegetative tissue and permit germination of the spores.
 |
| Figure 1 |
 |
| Figure 2 |
B. Fluorescence microscopy of GFP localizations
N. crassa strains were observed for GFP localization via fluorescence microscopy in Dr. Brian Shaw's lab. Five-second movies were recorded of growing hyphae (Figures 5 and 6 in Results).
C. Fungal Genetics Stock Center
Following is the link to the Fungal Genetics Stock Center website: http://www.fgsc.net/
N. crassa strains were observed for GFP localization via fluorescence microscopy in Dr. Brian Shaw's lab. Five-second movies were recorded of growing hyphae (Figures 5 and 6 in Results).
C. Fungal Genetics Stock Center
Following is the link to the Fungal Genetics Stock Center website: http://www.fgsc.net/
Founded in 1960 at Dartmouth College, the Fungal Genetics Stock Center (FGSC) is a resource available to fungal genetics
researchers that is funded mostly by a grant from the National Science Foundation of the United States of
America and to a lesser extent by payments made by researchers who use its
services. The FGSC was established to preserve strains of importance in genetics research. There were approximately 400 strains at the FGSC in 1960. Today the center maintains more than 19,000 Neurospora strains, a growing number of Neurospora knockouts, more than 2,000 Aspergillus strains and various
representatives
of other fungi. The FGSC also has cloned genes and gene libraries. Magnaporthe grisea strains and the molecular tools to work with them were added in 2001. Nearly 50,000
Magnaporthe knockout mutants were accepted in 2003 and 2004. The FGSC began to distribute arrayed sets of
knockout mutants of Cryptococcus and Candida mutants in 2005. For more information, see McCluskey, K., Wiest, A. and Plamann, M. 2010. The
Fungal Genetics Stock Center: a repository for 50 years of fungal genetics
research. J Biosci.
35(1):119-26.
D. Observation of fungal structures of Chlorophyllum sp.
Co-laboratory instructor Dr. Brian Shaw brought in a mushroom (Chlorophyllum sp.) he collected from his residence (Figure 3 below). We were instructed to remove a portion of the mushroom's hymenium (on the vertical face of the gill) (Figure 4 below) and microscopically examine it for basidia, basidiospores, hyphal clamp connections and Buller's drop. I used a scalpel to remove one of the mushroom gills and to cut a small section of tissue from the stem. I then placed the gill and stem section in drops of water on microscope slides and covered them with cover slips. I observed fungal structures using a compound microscope. Photographs were taken through the eyepiece of the microscope using a digital camera.
Co-laboratory instructor Dr. Brian Shaw brought in a mushroom (Chlorophyllum sp.) he collected from his residence (Figure 3 below). We were instructed to remove a portion of the mushroom's hymenium (on the vertical face of the gill) (Figure 4 below) and microscopically examine it for basidia, basidiospores, hyphal clamp connections and Buller's drop. I used a scalpel to remove one of the mushroom gills and to cut a small section of tissue from the stem. I then placed the gill and stem section in drops of water on microscope slides and covered them with cover slips. I observed fungal structures using a compound microscope. Photographs were taken through the eyepiece of the microscope using a digital camera.
 |
| Figure 3 |
 |
| Figure 4 |
Figure 5
Figure 6
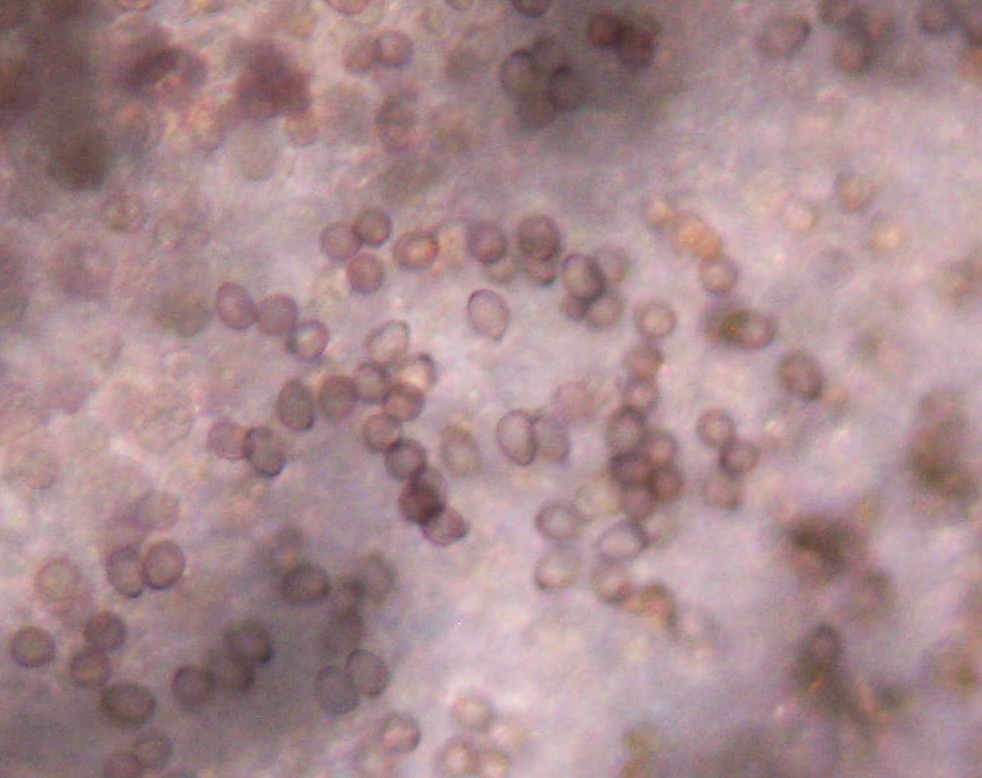 |
| Basidiospores at 40X. Photograph cropped and enlarged to show detail. |
 |
| Basidiospores still attached to basidia (40X). Buller's drops may be present in this sample. Notice two basidiospores seem to be leaning to the side. Photograph cropped and enlarged to show detail. |
 |
| Basidium with four basidiospores still attached (40X). Photograph cropped and enlarged to show detail. |
 |
| The stem of the mushroom is comprised of strands of hyphae (40X). No clamp connections were observed in this sample. Photograph cropped and enlarged to show detail. |
Discussion
Picking ascospores can be frustrating for a novice (ME!). I do not know whether the ascospores from the N. crassa SMRP10 x CSP-1-GFP cross germinated, as the slants were not returned to a subsequent laboratory for observation.
Picking ascospores can be frustrating for a novice (ME!). I do not know whether the ascospores from the N. crassa SMRP10 x CSP-1-GFP cross germinated, as the slants were not returned to a subsequent laboratory for observation.
Below is a figure to define Buller's drop and its role in dispersal of basidiospores.

Figure 7. Ballistospore discharge in Basidiomycetes. (A) Section of a typical
mushroom cap showing the gills and the location of the spore-bearing basidia
(insert). The approximate trajectory of the spore is shown as a broken line.
(B) A typical basidium with four spores. (C) Structure of the lower half of
the spore (based on McLaughlin et al. 1985).
(D) Spore ejection in Auricularia auricula. In this species, spores
are borne singly on the sporogenic surfaces. (E) Diagrammatic representation
of the ejection in D. http://jeb.biologists.org/content/212/17/2835.full























































.jpg)










{kind=link}